環形動物(かんけいどうぶつ)とは、環形動物門(学名: Annelida)に属する動物の総称である。多くが原則として体節制をもち、体は環状の柔らかい体節に分かれている蠕虫状の動物である。
環帯類(ミミズとヒル)、多毛類(ゴカイなど)を含むほか、有鬚動物(ヒゲムシやハオリムシ)、ユムシ動物、星口動物(ホシムシ)、スイクチムシ類を含む事が分子系統解析から分かっている[1][2][3][4]。多毛類は非単系統群であり[4]、ユムシやホシムシなどは二次的に体節構造を失ったと考えられている。
学名の由来は「環 annelum」である[5]。
体制
外形は蠕虫様の三胚葉性の左右相称動物で、大きな裂体腔を持つ[6]。
有鬚動物やユムシ動物、星口動物といったグループではほかの環形動物のもつ体節性や剛毛、消化器官といったボディプランの重要な構成要素の何れかを、進化の過程で退化・消失している[4]。そのため特に示さない場合、本節では環形動物とは便宜上それらを除いた環形動物を指す。また、便宜上多系統であることがわかっている多毛類という分類を用いる。
体節性
口前葉
囲口節
O 口
体節の増加部分
尾節
O 肛門
環形動物は各体節の機能的分化を伴う多体節性である[4]。多毛類では基本構造のよく似た多数の環状の体節が1列に連なっており、体内では体節と体節が隔壁 (septa)で区切られる[5]。最も先端にある口前葉 (prostomium)は目や感覚手、副触手、感覚器といった付属器官を具える[7][6]。それに続く体節は囲口節 (peristomium)で、感触糸などをもつ[5][6]。口前葉および囲口節に加え数体節が変形して頭部を形成する[5]。成体の胴部には体節が続き、胴部は胸部と腹部に分化することもある[5]。末端には肛門があり、肛触糸を持つことがある尾節 (pygidium)がある[5][6]。尾節付近で体節が増加する[6]。ヒル類では体の前後端にそれぞれ吸盤を持つことで体節が著しく変形している[5]。スイクチムシ類には不完全な体節がある[6]。
有鬚動物では前体 (protosoma)、中体 (mesosoma)、胴部 (metasoma)、後体 (opistosoma)の4域に分けられ、非常に短い後体にのみ剛毛を備えた複数の体節(剛毛節)が存在する[4][7]。前体と中体は「(広義の)前体 forepart」と呼ばれることもある[7]。そのうちハオリムシ類では前体は殻蓋部 (obturaculum)、中体はハオリ部 (vestimentum)となり、胴部は栄養体 (trophosoma)と呼ばれる[7]。後体はヒゲムシ類では複数の体節、ハオリムシ類では5-100体節からなる[7]。
ユムシ動物では幼生には表面的な体節性が現れるが、変態すると消失し、成体の体幹は無体節となる[4]。ただし腹神経索には痕跡的な体節性がある[4]。星口動物は体幹は一生を通じて無体節で、幼生時のみ腹神経索には痕跡的な体節性がある[4]。
剛毛
環形動物は普通キチン質の剛毛を備える[4]。多毛類では多数の剛毛が疣足に生じるのに対し、貧毛類では疣足を持たず少数の剛毛が直接体表に生じる[5]。ヒル類ではケビル Acanthobdella peledinaなどを除き剛毛を欠く。有鬚動物は後体にのみ剛毛を具える[7]。ユムシ動物では口の直後に腹剛毛、肛門を囲んで尾剛毛を持つものもいる[8]。星口動物は剛毛を一切持たない[4]。
神経系
環形動物の神経系は頭部背側の発達した中枢神経節(頭神経節[4])、神経環、腹側神経索からなる[6]。多毛類では多くの場合各体節にある1対の神経節が連絡し腹側神経索を形成するが、環帯類では中央で1本に融合している[5]。ハオリムシ類では体先端部の2本の神経索がハオリ部後部から1本に融合している[5]。ユムシ動物では1本の腹神経索が走り、その前端は2つに分岐して吻の周縁部を環状に1周するが、脳や神経節は形成されない[8]。星口動物の神経系は脳・腹神経索およびそれらをつなぐ食道神経環からなる[9]。脳は小型で咽頭背壁に接し、腹神経索は腹正中を走り1本で分節しない[9]。
吻
多毛類では、浮遊性のウキゴカイ類およびサシバゴカイ類、匍匐性のゴカイ類、チロリ類、シロガネゴカイ類などの捕食性の種では肉質の小突起を多数もつ外翻性の吻 (proboscis)を有することが多い[5]。
ユムシ動物は前背端から吻 (proboscis)が突出し、体幹中へは引き込まれない[8]。星口動物では体幹の前端に自由に出し入れできる陥入吻 (introvert)を持つ[9]。
発生
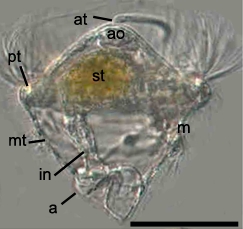 Pomatoceros lamarckiiのトロコフォア幼生。
Pomatoceros lamarckiiのトロコフォア幼生。
a: 肛門 anus, ao: 頂器官 apical organ, at: 頂毛の束 tuft of apical cilia, m: 口 mouth, mt: 口後繊毛環 metatroch, pt: 口前繊毛環 prototroch, st: 胃 stomach, in: 腸管 intestine
多毛類の多く、ハオリムシ類、ユムシ動物、星口動物では軟体動物と同様、螺旋卵割を行いトロコフォア幼生を経て発生が進行する[5][7][9][8]。シリス類、ウズマキゴカイ類では受精した卵が保育され、胚は定着準備の整った幼生として母体から放出される[5]。また、星口動物の中には直接発生をするものもいるが、トロコフォア幼生を経るものの中でもトロコフォアから成体になるもの、トロコフォアから卵黄栄養性ペラゴスフェラ幼生 (pelagosphera larva)になり成体へと変態するもの、トロコフォアから摂餌性ペラゴスフェラ幼生になって長期間遊泳するものもいる[9]。
トロコフォア幼生はコマのような形をしており、原口に由来する口をはさんで口前繊毛環 (prototroch)と口後繊毛環 (metatroch)、肛門の直上に端繊毛環 (telotroch)と、3つの繊毛環を持つ[5]。後期トロコフォア幼生の口前繊毛環より前方を上体 (episphere)と呼び、この部分が成体の口前葉に当たる[5]。また、成体の尾節に当たる部分のことを尾節部と呼ぶ[5]。
環帯類は限られた体節にのみ生殖巣が発達し、体前部の数節を覆って受精卵の卵包を分泌する環帯を持つ[5]。雌性生殖器と雄性生殖器を同時に用いて交接し、精包を交換する。受精卵は卵包に包まれて発生が進行し、幼生を経ずに幼虫が孵化する[5]。
系統関係
従来考えられていた、星口動物やユムシ動物を含む Rouse and Fauchald (1997)による形態に基づいた系統関係は次の通りである[1][10]。
分子系統解析の結果、多毛類やその中の分類群は解体され、次のような系統樹が描かれている[1]。太字で示したもの以外は全てかつての多毛類である。
なお、科より高次の分類群が示されていない部分はそれらをまとめた高次分類群は未だ命名されていないためである[1]。
かつて原始環虫類と呼ばれたグループの系統的位置については不明な点が多いが、一部は定在類や遊在類に含まれるとする分子系統解析の結果も得られている[11][12]。
脚注
注釈
- ^ 左上:ユムシ類の1種 Arhynchite hayaoi
右上:シボグリヌム科の1種 Lamellibrachia luymesi
左中:ゴカイ類の1種 Hesiocaeca methanicola
中央:スイクチムシ類の1種 Notopharyngoides aruensis
右中:ミミズ類の1種シマミミズ Eisenia fetida
左下:ケヤリムシ類の1種 Sabellastarte spectabilis
右下:ヒル類の1種 Haemadipsa zeylanica
出典
- ^ a b c d Weigert, Anne; Bleidorn, Christoph (2016). “Current status of annelid phylogeny”. Org Divers Evol 16: 345-362. doi:10.1007/s13127-016-0265-7.
- ^ Struck, Torsten H.; Paul, Christiane; Hill, Natascha; Hartmann, Stefanie; Hösel, Christoph; Kube, Michael; Lieb, Bernhard; Meyer, Achim et al. (2011-03). “Phylogenomic analyses unravel annelid evolution” (英語). Nature 471 (7336): 95–98. doi:10.1038/nature09864. ISSN 1476-4687. https://www.nature.com/articles/nature09864.
- ^ Weigert, Anne; Bleidorn, Christoph (2016-06). “Current status of annelid phylogeny” (英語). Organisms Diversity & Evolution 16 (2): 345–362. doi:10.1007/s13127-016-0265-7. ISSN 1439-6092. http://link.springer.com/10.1007/s13127-016-0265-7.
- ^ a b c d e f g h i j k 日本動物学会 2018 pp.70-71
- ^ a b c d e f g h i j k l m n o p q r 三浦・白山 2000, pp.203-211
- ^ a b c d e f g 藤田 2010, p.142-148
- ^ a b c d e f g 三浦・白山 2000, pp.199-202
- ^ a b c d 西川 2000-23, pp.196-198
- ^ a b c d e 西川 2000-22, pp.193-195
- ^ 和名は『岩波生物学辞典 第5版』(2013), pp.1584-1586に基づく
- ^ Andrade, Sónia C.S.; Novo, Marta; Kawauchi, Gisele Y.; Worsaae, Katrine; Pleijel, Fredrik; Giribet, Gonzalo; Rouse, Greg W. (2015-11). “Articulating “Archiannelids”: Phylogenomics and Annelid Relationships, with Emphasis on Meiofaunal Taxa” (英語). Molecular Biology and Evolution 32 (11): 2860–2875. doi:10.1093/molbev/msv157. ISSN 0737-4038. https://academic.oup.com/mbe/article-lookup/doi/10.1093/molbev/msv157.
- ^ Struck, Torsten Hugo; Golombek, Anja; Weigert, Anne; Franke, Franziska Anni; Westheide, Wilfried; Purschke, Günter; Bleidorn, Christoph; Halanych, Kenneth Michael (2015-08). “The Evolution of Annelids Reveals Two Adaptive Routes to the Interstitial Realm” (英語). Current Biology 25 (15): 1993–1999. doi:10.1016/j.cub.2015.06.007. https://linkinghub.elsevier.com/retrieve/pii/S0960982215006727.
参考文献
- 白山義久編、岩槻邦男・馬渡峻輔 監修『無脊椎動物の多様性と系統 : 節足動物を除く』裳華房〈バイオディバーシティ・シリーズ〉、2000年。ISBN 4-7853-5828-9。
- 西川輝昭 著「22. 星口動物門 Phylum SIPUNCULA」、白山義久編 編『無脊椎動物の多様性と系統 : 節足動物を除く』岩槻邦男・馬渡峻輔 監修、裳華房〈バイオディバーシティ・シリーズ〉、2000年、193-195頁。ISBN 4-7853-5828-9。
- 西川輝昭 著「23. ユムシ動物門 Phylum ECHIURA」、白山義久編 編『無脊椎動物の多様性と系統 : 節足動物を除く』岩槻邦男・馬渡峻輔 監修、裳華房〈バイオディバーシティ・シリーズ〉、2000年、196-198頁。ISBN 4-7853-5828-9。
- 三浦知之・白山義久 著「24. 有鬚動物門 Phylum POGONOPHORA」、白山義久編 編『無脊椎動物の多様性と系統 : 節足動物を除く』岩槻邦男・馬渡峻輔 監修、裳華房〈バイオディバーシティ・シリーズ〉、2000年、199-202頁。ISBN 4-7853-5828-9。
- 三浦知之・白山義久 著「25. 環形動物門 Phylum ANNELIDA」、白山義久編 編『無脊椎動物の多様性と系統 : 節足動物を除く』岩槻邦男・馬渡峻輔 監修、裳華房〈バイオディバーシティ・シリーズ〉、2000年、203-211頁。ISBN 4-7853-5828-9。
- 巌佐庸・倉谷滋・斎藤成也・塚谷裕一『岩波生物学辞典 第5版』岩波書店、2013年2月26日。ISBN 9784000803144。
- 公益社団法人日本動物学会『動物学の百科事典』丸善出版、2018年9月28日。ISBN 978-4621303092。
- 藤田敏彦、駒崎伸二 著、太田次郎、赤坂甲治、浅島誠、長田敏行 編『動物の系統分類と進化』裳華房〈新・生命科学シリーズ〉、2010年4月28日。ISBN 978-4785358426。
関連項目
外部リンク
ウィキメディア・コモンズには、
環形動物門に関連するカテゴリがあります。
ウィキスピーシーズに
環形動物門に関する情報があります。


