Een reeks codons in het messenger-RNA (mRNA). Elk codon bestaat uit drie nucleotiden, en de volgorde daarvan bepaalt de volgorde van aminozuren, de bouwstenen van een eiwit. In het mRNA worden de nucleotiden aangegeven met de letters A, U, G en C.
De genetische code beschrijft hoe de informatie in het genetisch materiaal (DNA of RNA) wordt gelezen om een eiwit te vormen. De genetische code werd ontrafeld in de jaren 1960, enkele jaren na de ontdekking van de DNA-helix. Het verklaart hoe de vierletterige code van het DNA vertaald kan worden naar de twintig verschillende aminozuren, de bouwstenen van eiwitten. Op enkele uitzonderingen na is de genetische code universeel geldig bij alle vormen van leven.
Stukken DNA die coderen voor eiwitten (genen), worden in levende cellen overgeschreven naar een enkelstrengs RNA-molecuul. Dit molecuul, het messenger-RNA, is in wezen een kopie van het DNA en draagt instructies voor de aanmaak van een eiwit. In het ribosoom worden aminozuren aan elkaar gekoppeld in een volgorde die bepaald wordt door de nucleotidevolgorde in het mRNA. Het ribosoom leest daarbij steeds drie nucleotiden af van het mRNA, elk overeenkomend met een specifiek aminozuur. Dit proces wordt translatie genoemd.
De drie opeenvolgende nucleotiden, noemt men een codon. Codons specificeren welke aminozuren er aan de eiwitketen toegevoegd worden, en daarnaast bepalen ze de start- en stopsignalen van het translatieproces. Meestal doelt men met de term codon op de nucleotiden zoals die voorkomen in messenger-RNA: adenine, cytosine, guanine en uracil. Het codon 'ACU' correspondeert bijvoorbeeld met het aminozuur threonine, en 'CGG' met arginine. De genetische code kan samengevat worden in een simpele codontabel van 64 elementen.
Geschiedenis
Het codonkompas, een manier om de genetische code visueel weer te geven. Nucleotiden van het mRNA gekleurd, en de resulterende aminozuren aan de buitenrand.
Al lang voor de ontdekking van de moleculaire basis van erfelijkheid was duidelijk dat overerfbare factoren (genen) instructies moeten bevatten voor de aanmaak van eiwitten. Eiwitten staan aan de basis van alle celactiviteit, en hun biologische functie wordt bepaald door hun specifieke ruimtelijke structuur. Deze structuur is afhankelijk van de volgorde van aminozuren waaruit het eiwit bestaat. De lineaire sequentie van nucleotiden in het DNA moet dus op een bepaalde manier te vertalen zijn naar een sequentie van aminozuren. Hoe het vierletterige DNA-alfabet overeenkomt met het 20-letterige aminozuuralfabet – de genetische code – werd ongeveer een decennium na de ontdekking van de DNA-helix ontrafeld.[1]
De Russisch-Amerikaanse wetenschapper George Gamow was een van de eersten die een werkbaar schema opstelde voor de translatie van DNA naar eiwit. In samenwerking met Francis Crick kwam hij op het idee dat drie opeenvolgende basen (een triplet) gebruikt konden worden om voor twintig proteïnogene aminozuren te coderen.[2] Hij constateerde dat 43 = 64 permutaties gereduceerd kon worden tot 20 combinaties, als de volgorde van de basen niet uitmaakt. Later werd duidelijk dat de volgorde van letters binnen een triplet wel degelijk uitmaakt: Gamow's model bleek niet geheel correct. Men kwam er later achter dat er sprake is van redundantie is in de genetische code; sommige nucleotide-tripletten coderen voor hetzelfde aminozuur. Dit inzicht was een belangrijke stap in de ontrafeling van de genetische code.
Ontrafeling van codons
Het bewijs dat de genetische code inderdaad gevormd wordt groepjes van drie opeenvolgende nucleotiden, werd geleverd door Francis Crick en Sydney Brenner.[a]Marshall Niremberg en zijn collega's slaagden er in 1961 in het eerste codon te ontcijferen.[3] Ze gebruikten een synthetisch RNA-molecuul dat alleen bestond uit uracil (dus UUUUU...) en ontdekten dat dit getransleerd werd tot een polypeptide dat uitsluitend bestond uit het aminozuur fenylalanine. Daaruit concludeerden ze dat het codon UUU moest coderen voor het aminozuur fenylalanine.
Dit experiment werd doorgezet door onder meer Severo Ochoa, die ontdekte dat een poly-adenine RNA-molecuul (AAAAA...) getransleerd werd tot een polypeptide van alleen lysine, en poly-cytosine RNA (CCCCC...) getransleerd werd naar een polypeptide met alleen proline. Hieruit leidde hij af dat het codon AAA voor het aminozuur lysine codeert, en CCC voor proline. Door gebruik te maken van copolymeren werden in de loop van de jaren 1960 alle andere codons geïdentificeerd. Niremberg was hierin een vooraanstaand onderzoeker.[4]
Rond deze tijd werd ook de rol van transfer-RNA (tRNA) in het translatieproces opgehelderd: dit molecuul fungeerde als een adaptor die de verbinding vormde tussen aminozuren en nucleotiden in het RNA. Onderzoekers Har Gobind Khorana, Robert W. Holley en Nirenberg wisten een groot deel van de puzzel op te lossen, en hun werk werd bekroond met de Nobelprijs voor Fysiologie of Geneeskunde in 1968.[5]
Het leesraam van twee deels overlappende genen, in het mitochondriaal DNA. Het gen ATP8 (rood) wordt gelezen vanaf de +1 base, en stopt bij het stopcodon 'TAG' (rode stip). Een ander gen, ATP6 (blauw), begint bij base +3. Dit is dus een ander leesraam, en leidt tot een totaal ander eiwitproduct. Merk op dat de codons altijd in de richting 5' naar 3' worden gelezen.
Hoewel men DNA- of RNA-sequenties vaak weergeeft zonder spaties tussen de codons, worden ze in levende cellen altijd gelezen als aparte, drieletterige woorden.[1] Deze vaste opdeling van een nucleotidesequentie in groepjes van drie noemt men in de moleculaire biologie het leesraam.
De sequentie 5'-AAATGAACG-3' bevat bijvoorbeeld, indien gelezen vanaf de eerste positie, de codons AAA, TGA en ACG; als het gelezen wordt vanaf de tweede positie bevat het de codons AAT en GAA; en als het vanaf de derde positie wordt gelezen, bevat het de codons ATG en AAC. Een sequentie kan dus gelezen worden in drie mogelijke leesramen. Afhankelijk van hoe je het leesraam kiest, ontstaat er een compleet andere aminozuurvolgorde. Eiwitcoderende genen beginnen altijd met een startcodon, en lopen tot een stopcodon. Het hele stuk, vanaf start tot stop, noemt men ook wel een open leesraam.
Start- en stopcodons
Niet elk van de 64 codons in de codontabel codeert voor een aminozuur. Er zijn drie codons (UAA, UAG en UGA)[b] die voor géén aminozuur coderen: dit zijn de stopsignalen die het einde markeren van het translatieproces. Wanneer een ribosoom een stopcodon tegenkomt, wordt er geen aminozuur meer ingebouwd in de polypeptideketen, omdat er geen tRNA-moleculen bestaan die een anticodon hebben tegen de stopcodons. Een eiwit genaamd de release factor herkent het stopcodon en zorgt ervoor dat de nieuwgevormde polypeptide loslaat van het ribosoom.[6]
De translatie begint met een startcodon. In eukaryoten is dit het codon 'AUG', wat eveneens codeert voor het aminozuur methionine. Bijna alle polypeptiden beginnen dus met het aminozuur methionine, hoewel dit eerste aminozuur er vaak in een later stadium wordt afgeknipt. In veel gevallen is het startcodon zelf niet voldoende om de translatie op gang te brengen. Het startcodon wordt in bacteriën als E. coli bijvoorbeeld vaak voorafgegaan door de Shine-Dalgarnosequentie, een stukje van zes nucleotiden dat nodig is voor de rekrutering van initiatiefactoren.
Redundantie en wiebelbasen
Een belangrijk principe van de genetische code is redundantie: er zijn verschillende codons die hetzelfde aminozuur specificeren. Deze redundantie komt voort uit het feit dat er meerdere verschillende tRNA-moleculen bestaan die hetzelfde aminozuur dragen, en uit het feit dat er tRNA-moleculen zijn die met meer dan één codon kunnen basenparen. Voor de herkenning van een tRNA-molecuul op een codon (via een anticodon), zijn voornamelijk de eerste twee posities binnen de triplet van belang. De derde positie is toleranter en laat toe dat hier een verkeerde basenparing of 'mismatch' optreedt.[1] Deze tolerante positie wordt de wiebelbase genoemd. De wiebelbase verklaart waarom synonieme codons die voor hetzelfde aminozuur coderen, vaak alleen verschillen in de derde nucleotide. Het aminozuur alanine wordt bijvoorbeeld gecodeerd door GCA, GCU, GCG en GCC.[6]
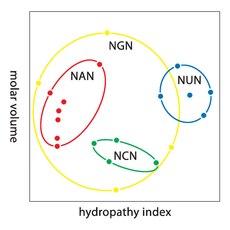
Groepering van codons op basis van hydropathiciteit en molaire grootte van de gecodeerde aminozuren. Codons met een U op de tweede positie (blauw) coderen bijvoorbeeld vooral voor middelgrote, hydrofobe aminozuren.
Een praktische consequentie van redundantie is dat mutaties die optreden in de wiebelpositie niet leiden tot een ander aminozuur: een stille mutatie.[6] In de gevallen dat er wel een ander aminozuur ontstaat, is dit aminozuur vaak in biochemische eigenschappen vergelijkbaar met het oorspronkelijke aminozuur. Het hydrofobe aminozuur fenylalanine (codons UUC en UUU), zal bijvoorbeeld veranderen in het eveneens hydrofobe aminozuur leucine als de derde base muteert (UUA en UUG). Dit zal de eiwitfunctie waarschijnlijk niet ingrijpend veranderen. Op deze manier kan de functie van belangrijke eiwitten in de loop van de evolutie behouden worden hoewel er soms puntmutaties optreden.
Wiskundige analyses laten zien dat de genetische code zeer sterk ingericht is op behoud van hoe hydrofoob of hydrofiel een aminozuur is. Wanneer men een singulierewaardenontbinding (4 nucleotiden × 3 posities) loslaat op de sequenties van codons, levert dit een opmerkelijke correlatie op (C = 0,95) voor het voorspellen van de hydrofobiciteit van het gecodeerde aminozuur.[7]
Codongebruik
Hoewel er meerdere codons zijn die voor één aminozuur coderen, wordt niet elk codon even vaak gebruikt tijdens de translatie. De frequentie waarmee een codon gekozen wordt, verschilt vaak van organisme tot organisme, en wordt aangeduid met het begrip codon usage bias.[8] De bacterie E. coli maakt bijvoorbeeld het meest gebruik van het codon CCG om proline te transleren, terwijl dit bij mensen het minst gebruikte codon voor proline is. Deze bias is een gevolg van verschillen in de relatieve aanwezigheid van tRNA-moleculen binnen de cel. Codon usage bias is van belang in de genetische technologie en biotechnologie: om de expressie van een kunstmatig gemaakt gen hoog te krijgen, moeten de codons voor het gastheerorganisme worden geoptimaliseerd.[8]
Lange tijd werd aangenomen dat de genetische code universeel is; een gegeven codon zou in elk organisme voor hetzelfde aminozuur coderen. Deze aanname is grotendeels waar: de genetische code is van toepassing bij alle drie de domeinen van het leven. Dit feit levert een belangrijk argument voor de gemeenschappelijke afstamming van alle levensvormen. Er zijn echter een paar uitzonderingen – variaties – op de genetische code ontdekt.[9] De eencellige schimmel Candida albicans transleert het codon 'CUG' bijvoorbeeld als serine, terwijl bijna alle andere organismen dit codon als leucine transleren. Bij sommige ciliaten (eencellige eukaryoten) werken de drie conventionele stopcodons anders; deze coderen namelijk gewoon voor aminozuren, en het eind van de translatie wordt gesignaleerd door het 3'-eind van het mRNA.[9]
In mitochondriën van uiteenlopende soorten geldt ook een sterk variërende genetische code. In de mitochondriën van zoogdieren wordt het codon 'AUA' bijvoorbeeld getransleerd als methionine, in plaats van 'AUG'. Mitochondriën hebben hun eigen genetisch materiaal en coderen hun eigen translatiemachinerie. Dergelijke variaties laten zien dat de genetische code niet zonder meer onwrikbaar vastgelegd is, maar evolutionaire veranderingen kan ondergaan.[10]
↑In dit experiment werden kleine deleties gemaakt in een willekeurig gen (rIIB) van de T4-bacteriofaag. Wanneer één of twee basenparen werden verwijderd uit de coderende regio, was het genproduct verstoord door een leesraamverschuiving. Wanneer er drie baseparen werden verwijderd, kon nog steeds een functioneel eiwit worden getransleerd, omdat slechts één aminozuur uit het eiwit ontbrak.
↑De drie stopcodons hebben namen: UAG heet amber, UGA heet opal en UAA heet ochre.
↑(en) Szymanski M, Barciszewski J. (2017). The path to the genetic code. Biochimica et Biophysica Acta: 2674-2679. DOI: 10.1016/j.bbagen.2017.07.009.
↑(en) Yanofsky C. (2007). Establishing the Triplet Nature of the Genetic Code. Cell128 (5): 815-818. DOI: 10.1016/j.cell.2007.02.029.
↑(en) Nirenberg M, Leder P, Bernfield M, Brimacombe R, Trupin J, Rottman F, O'Neal C (1965). RNA codewords and protein synthesis, VII. On the general nature of the RNA code. Proceedings of the National Academy of Sciences of the United States of America53 (5): 1161–8. DOI: 10.1073/pnas.53.5.1161.